Raphinae
 | ||
|---|---|---|
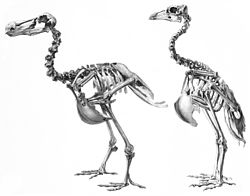 Comparación entre los esqueletos del dodo y del solitario de Rodrigues. | ||
| Taxonomía | ||
| Reino: | Animalia | |
| Filo: | Chordata | |
| Clase: | Aves | |
| Orden: | Columbiformes | |
| Familia: | Columbidae | |
| Subfamilia: | †Raphinae Wetmore, 1930 | |
| Distribución | ||
 Antiguo rango geográfico (en rojo). | ||
| Especies | ||
| ||
| Sinonimia | ||
Lista Didi Bonaparte, 1857 Raphi Ridgeway, 1916 Didiadae Swainson, 1836 Dididae Swainson, 1837 Didusidae Lesson, 1842 Raphidae Poche, 1904 (no disponible)[nota 1][1] Raphidae Wetmore, 1930 Didinae Strickland & Melville, 1848[2] Pezophapidae Hachisuka, 1953 Pezophabidae (Hachisuka, 1953) Illiger, 1811 (no disponible)[nota 2][3][1] Raphini Verheyen, 1957 Pezophabini Verheyen, 1957 | ||
[editar datos en Wikidata] | ||
Raphinae es un clado de aves no voladoras extintas, llamadas antiguamente didinos,[nota 3] las cuales habitaron las Islas Mascareñas de Mauricio y Rodrigues. Este grupo incluye al dodo (Raphus cucullatus) y al solitario de Rodrígues (Pezophaps solitaria). Históricamente, muchos grupos diferentes han sido nombrados tanto para el dodo como para el solitario de Rodrígues, aunque no todos los agrupan a ambos. Más recientemente, se ha considerado que ambas aves deben ser clasificadas en Columbidae, con frecuencia bajo la subfamilia Raphinae. La primera persona en sugerir una cercana afinidad con las palomas fue Johannes Theodor Reinhardt, cuyas opiniones fueron apoyadas por Hugh Edwin Strickland y Alexander Gordon Melville.
Los análisis realizados con ADN extraído del dodo y el solitario de Rodrígues, junto al de 37 especies de palomas han encontrado que los rafinos se incluyen dentro de Columbidae. Sorprendentemente, los rafinos no son los colúmbidos más primitivos, en realidad se agrupan junto con la paloma de Nicobar como su pariente vivo más cercano, siendo otros parientes cercanos las guras y la paloma manumea. Un tercer rafino, Raphus solitarius, es considerado ahora como un ibis del género Threskiornis.
Una fecha común para la extinción del dodo es 1662, aunque hay posibles avistamientos tan tardíos como 1688. El último avistamiento con una descripción fue en 1662, pero un análisis estadístico realizado por Roberts y Solow encontró que la extinción del dodo debió producirse en 1693. El solitario de Rodrígues desapareció más tarde que el dodo. La IUCN usa como fecha de su extinción el año de 1778, aunque es más probable que la fecha este entre las décadas de 1750 a 1760. Ambas aves finalmente se extinguieron a raíz de la caza y la introducción de especies de mamíferos no-nativos tras la colonización del hombre en el siglo XVII.
Taxonomía
Este clado es parte del orden Columbiformes y contiene a los géneros monotípicos Pezophaps y Raphus. El primero contiene a la especie Pezophaps solitaria (el solitario de Rodrigues), y el último al dodo, Raphus cucullatus. Estas aves alcanzaron un gran tamaño como resultado del aislamiento en islas que estaban libres de depredadores, siguiendo la regla de Foster.[4]
Historia de la clasificación
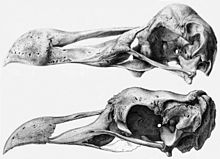
Históricamente, el dodo fue asignando al género Didus, ahora un sinónimo más moderno de Raphus.[3] En 1848, se incluyó a una nueva especie dentro de este género obsoleto, D. nazarenus por Hugh Edwin Strickland y Alexander Gordon Melville. Para alojar a futuras nuevas especies, así como a las especies conocidas en su época, Strickland y Melville nombraron a la subfamilia Didinae.[2] En 1893 tres especies fueron asignadas al grupo, Pezophaps solitarius, Didus ineptus, y la posible especie Didus borbonicus.[5] Hoy en día, solo se reconocen dos especies de rafinos, siendo Didus ineptus un sinónimo más moderno subjetivo de Raphus cucullatus; Didus? borbonicus es clasificado como el ibis Threskiornis solitarius;[3] y Didus nazarenus ha sido identificado como sinónimo de Pezophaps solitarius.[6]
Un suborden nombrado en 1893 por Sharpe, Didi, fue definido como un grupo que incluía solo a aves grandes que eran próximas a las Columbidae, de las islas Mascareñas de Mauricio, Reunión y Rodríguez. Los rasgos que agrupaban a Didi con Columbidae eran el ángulo de la mandíbula y el gancho en el extremo del pico.[5]
En 1811, Johann Karl Wilhelm Illiger creó una nueva familia para el género Didus. Él la denominó Inepti, y en esta incluyó solo a Didus ineptus, ahora un sinónimo de Raphus cucullatus. Illiger concluyó que el dodo estaba emparentado con los avestruces y ñandúes, por lo que situó a Inepti en el orden Rasores, como una familia cercana a los Gallinacei, Epollicati (un grupo inválido que incluía a los géneros Turnix y Syrrhaptes[7]), Columbini y a Crypturi.[3]
En 1842, Johannes Theodor Reinhardt propuso que estas eran palomas terrestres, basándose en estudios de un cráneo de dodo que él redescubrió en la colección real danesa en Copenhague.[8] Esta idea fue ridiculizada, pero más tarde recibió apoyo de Strickland y Melville, quienes sugirieron un ascendente común para el solitario y el dodo en 1848, tras diseccionar el único espécimen conocido de dodo con tejidos blandos y compararlo con los pocos restos entonces disponibles del solitario.[9] Strickland estableció que aunque no eran idénticas, estas aves compartían muchas características distintivas en los huesos de las patas, rasgos que por lo demás solo se conocían en las palomas.[2]
Los rafinos son a veces separados como una familia distinta denominada Raphidae, y sus afinidades fueron por mucho tiempo inciertas. Fueron situadas en principio entre las ratites debido a sus peculiares apomorfias relacionadas con la inhabilidad de volar, y se ha sugerido también una relación con los Rallidae. Sin embargo, los datos osteológicos y moleculares concuerdan en que lo más apropiado es colocarlos dentro de Columbidae.[10] Se han propuesto muchas afinidades diferentes para el dodo, incluyendo que fuera un pequeño avestruz, un rállido, un albatros o un buitre.[11]
Filogenia
La comparación de las secuencias de ADN del citocromo b mitocondrial y el rRNA 12S aislados de un tarso de un dodo y un fémur del solitario de Rodrigues confirmaron su cercana relación y su clasificación dentro de Columbidae.[12] Se interpretó que la evidencia genética mostraba que la paloma de Nicobar del Sureste asiático era su pariente vivo más cercano, seguida por las guras de Nueva Guinea y la paloma manumea superficialmente parecida al dodo, procedente de Samoa.[13] El nombre de género de esta última es Didunculus ("pequeño dodo"), y fue llamada "dodlet" ("dodito") por Richard Owen.[14] El siguiente cladograma, basado en Shapiro y colaboradores (2002), muestra la cercana relación del dodo con el resto de las Columbidae, un clado que consiste en general de especies terrestres endémicas de islas.[12] En 2014, el ADN del único espécimen conocido de la recientemente extinta paloma verde manchada (Caloenas maculata) fue analizado, y se encontró que era un pariente cercano de la paloma de Nicobar, y por tanto también del dodo y el solitario de Rodrigues.[15]
| |||||||||||||||||||||||||||||||

Un cladograma parecido fue publicado en 2007, aunque invirtiendo la posición de Goura y Didunculus e incluyendo a la paloma faisán y a la paloma terrestre de pico grueso en la base del clado.[16] Basándose en la evidencia molecular y de comportamientos, Jolyon C. Parish propuso que el dodo y el solitario de Rodrigues debería ser situados en la subfamilia Gourinae junto con las palomas Goura y otras, de acuerdo con la evidencia genética.[17]
Por muchos años el dodo y el solitario de Rodrigues fueron situados en su propia familia, Raphidae (anteriormente Dididae), debido a que sus relaciones precisas con otras palomas no podían ser resueltas. Cada una fue situada también en su propia familia monotípica (Raphidae y Pezophapidae, respectivamente), ya que se pensaba que habían evolucionado sus rasgos similares de manera independiente.[18] Los datos osteológicos y moleculares han llevado posteriormente a la disolución de la familia Raphidae, y el dodo y el solitario han sido colocados en su propia subfamilia, Raphinae,[nota 4] en la familia Columbidae.[19]
El "solitario de Reunión", por largo tiempo considerado como una tercera ave de este grupo, ha sido reclasificada como un ibis; es conocida desde entonces como Threskiornis solitarius.[20]
Divergencia
El estudio de 2002 indicó que los ancestros del solitario de Rodrigues y el dodo divergieron alrededor del límite entre el Paleógeno y el Neógeno. Las islas Mascareñas (Mauricio, Reunión y Rodrigues), son de origen volcánico y tienen menos de 10 millones de años de antigüedad. Por lo tanto, los ancestros de ambas aves probablemente eran aún capaces de volar por un periodo considerable tras la separación de sus linajes evolutivos.[21] La carencia de mamíferos herbívoros que compitieran por los recursos en estas islas permitió que el solitario y el dodo alcanzaran grandes tamaños.[22] El ADN obtenido del espécimen de Oxford está degradado, y no se ha extraído ADN utilizable de los restos subfósiles, por lo que la edad de la divergencia de estos con respecto a otras palomas aún necesita de verificación independiente.[23] El dodo perdió la habilidad de volar debido a la carencia de depredadores mamíferos en Mauricio.[24] Otra gran paloma no voladora, la paloma gigante de Viti Levu (Natunaornis gigoura), fue descrita en 2001 de material subfósil de Fiyi. Era solo levemente más pequeña que el dodo y el solitario de Rodrigues, y también se cree que estaba relacionada con las palomas guras.[25]
Se ha estimado que el grupo que contiene al dodo y al solitario de Rodrigues divergió de géneros como Goura hace unos 1.5 millones de años.[12] Sin embargo, esta estimación parece ser sumamente improbable. Otras estimaciones indican que los parientes de ambas especies llegaron a las islas hace 35 millones de años, cuando había un puente terrestre entre los bancos de Nazareth (Rodrigues) o Cargados Carajos y Mauricio se formaba.[3]
Descripción
La mayor parte de los rasgos del esqueleto que distinguen al dodo y al solitario de Rodrigues, su pariente más cercano de las demás palomas se ha atribuido a su inhabilidad de volar. Los elementos pélvicos eran más gruesos que los de las palomas voladoras para soportar su mayor peso, y la región pectoral y las pequeñas alas eran pedomórficos, lo que significa que no estaban bien desarrollados y retenían rasgos juveniles. El cráneo, el tronco y los elementos pélvicos eran peramórficos, lo que significa que cambiaban considerablemente con la edad. El dodo compartía varias otras características con el solitario de Rodrigues, como rasgos del cráneo, pelvis y el esternón, así como su gran tamaño. Difería en otros aspectos, como ser más robusto y corto que el solitario, tener un cráneo y pico más grandes, una bóveda craneana redondeada, y órbitas más pequeñas. El cuello y las patas del dodo eran proporcionalmente más cortas, y no poseía un equivalente de la protuberancia presente en las muñecas del solitario.[26]

Como no hay especímenes completos del dodo, su apariencia externa, como el plumaje y la coloración, son difíciles de determinar.[27] Las ilustraciones y crónicas escritas de los encuentros con el dodo entre su descubrimiento y su extinción (1598–1662) son las fuentes primarias sobre su apariencia externa.[28] De acuerdo con muchas representaciones, el dodo tenía un plumaje pardo o grisáceo, con plumas primarias claras y un mechón de plumas curvas claras en su cola. La cabeza era gris y desnuda, el pico verde, negro y amarillo, y las patas eran gruesas y amarillentas, con garras negras.[29] Restos subfósiles y remanentes de estas aves fueron llevados a Europa en el siglo XVII mostrando que eran aves muy grandes, de un metro de altura, y posiblemente pesando más de 23 kilogramos. Los pesos más altos han sido atribuidos a aves en cautiverio; se estima que en su medio natural habrían estado en el rango de 10.6 a 21.1 kilogramos.[30] Una estimación más tardía le da un peso promedio tan bajo como 10.2 kg.[31] Esto se ha cuestionado, y aún existe controversia al respecto.[32][33] Se ha sugerido que el peso dependía de la temporada, y que los individuos engordaban durante los períodos fríos, para perder peso cuando hiciera calor.[34] Esta ave era sexualmente dimórfica: los machos eran más grandes y tenía picos proporcionalmente más largos. El pico medía más de 23 centímetros de longitud y tenía una punta en forma de gancho.[26] Un estudio de las pocas plumas que quedan en la cabeza del espécimen de Oxford muestra que estas eran penáceas (plumas divididas con barbas y con cañón) en vez de ser plumáceas (vellosas) y más parecidas a las de otras palomas.[35]

El pico del solitario era levemente ganchudo, y su cuello y patas eran largos.[36] Un observador lo describió como del tamaño de un cisne.[37] El cráneo medía 170 milímetros de largo, aplanado en la parte superior con las partes delanteras y posteriores elevadas en dos bordes óseos estructurados con hueso poroso.[38] Una banda negra (una descripción contemporánea la describe como una "diadema"[nota 5] aparecía en su cabeza justo por detrás de la base del pico. El plumaje del solitario de Rodrigues fue descrito como gris y pardo. Las hembras eran de color más claro que los machos y tenían elevaciones de color claro en la parte inferior del cuello.[37] El dimorfismo sexual en tamaño en el solitario era quizás el más grande en cualquier ave neognata.[39] Un grupo, probablemente los machos, era considerablemente más grande que el otro, midiendo 90 centímetros de longitud y pesando más de 28 kilogramos, mientras que el grupo de individuos pequeños, presuntamente las hembras, medían solo 70 cm, y pesaban unos 17 kilogramos.[40] Esto constituye solo el 60% del peso de un individuo grande.[39] Su peso puede haber variado sustancialmente debido a los períodos de engorde, lo que implica que como en el dodo los individuos ganaban masa durante las temporadas frías y enflaquecían en las cálidas, bajando hasta solo 21 kilogramos en el sexo más grande y 13 kilogramos en el más pequeño.[26] Aunque las palomas macho generalmente son más grandes que las hembras, no hay evidencia directa de que los especímenes mayores sean el macho de la especie, y esto solo se supuso en los primeros documentos. Aunque el macho probablemente era el más grande, esto solo se puede confirmar mediante técnicas moleculares, y no solo con la morfología esquelética.[39]
Comportamiento

Poco se conoce sobre el comportamiento del dodo, debido a que muchas de las descripciones contemporáneas son muy breves.[30] Basándose en las estimaciones de peso, se ha sugerido que el macho podía alcanzar los 21 años de edad, y las hembras hasta 17.[26] Estudios de la fuerza del voladizo de los huesos de sus patas indican que podía correr muy rápidamente.[30] A diferencia del solitario de Rodrigues, no hay evidencia de que el dodo usara sus alas en combates con los de su propia especie. Aunque se han hallado algunos huesos de dodo con fracturas sanadas, tenía en comparación a su pariente músculos pectorales más débiles y alas más reducidas. El dodo en cambio puede haber usado su gran hocico ganchudo para las disputas territoriales. Dado que la isla de Mauricio recibía más lluvias y tenía un clima más estable que Rodrigues, probablemente había mucha menos necesidad para los machos de dodo de pelear por los territorios. Por lo tanto, el solitario sería el más agresivo de los dos.[39]
Las observaciones del solitario de Rodrigues en vida indican que eran animales sumamente territoriales. Probablemente establecían disputas al golpearse entre sí con sus alas; para ayudar a este propósito, ellos usaban los "puños" en sus muñecas.[41] Las fracturas de los huesos del ala también indican que eran usados en combate.[26] También se ha sugerido que estas fracturas pueden ser en cambio el resultado de una enfermedad ósea hereditaria en vez de ser heridas de combates.[42] Pero en todas las aves modernas que poseen protuberancias o espinas en los carpos, están son usadas como armas sin excepción. La menor cantidad de lluvias y las variaciones estacionales de la isla de Rodrigues pueden haber afectado la disponibilidad de recursos de la isla, por lo que el solitario tendría más razones para haber evolucionado un agresivo comportamiento territorial.[39] Varias crónicas atestiguan que estos se defendían además con una poderosa mordida.[37]
Extinción
La última especie en extinguirse de los rafinos, el solitario de Rodrigues, probablemente desapareció antes de 1778.[43] El dodo sobrevivió hasta 1662 o 1690.[44] El solitario de Rodrigues se extinguió debido a la introducción de gatos ferales y el exceso de caza por la población humana.[43] Aunque el dodo se extinguió antes, las causas de su extinción se relacionan también con la introducción de animales y la caza.[44]
Extinción del dodo
Como muchos animales que evolucionaron en aislamiento de grandes depredadores, el dodo carecía por completo de miedo a los humanos. Esta carencia de miedo y su incapacidad para volar convertían al dodo en una presa fácil para los marineros.[45] La población humana en Mauricio (en un área de 1,860 km²) nunca pasó de 50 personas en el siglo XVII, pero ellos llevaron a la introducción de varios animales tales como perros, cerdos, gatos, ratas y macacos cangrejeros, los cuales asaltaban los nidos de los dodos y competían con ellos por los limitados recursos alimenticios.[46] Al mismo tiempo, los humanos destruían el hábitat forestal del dodo. El impacto de estos animales introducidos, especialmente los cerdos y macacos, sobre la población de dodos es considerado como mucho más severo que el de la caza.[47] Las ratas no debieron de ser un gran problema para los dodos, ya que estaban acostumbrados a defenderse de los cangrejos terrestres locales.[48]
El último avistamiento definitivo de los dodos, en la isla de Amber en 1662, puede no haber sido realmente el último para esta especie.[49] Hubo un supuesto avistamiento de un dodo reportado en los registros de cacería de Isaac Johannes Lamotius en 1688. Los análisis estadísticos de estos registros realizados por Roberts y Solow dan una nueva fecha de extinción estimada en el año 1693, con un intervalo de confianza del 95% para 1688–1715. Los autores señalaron también que debido a que el último avistamiento antes de 1662 ocurrió en 1638, el dodo probablemente ya se había hecho muy raro en la década de 1660, y por lo tanto un reporte cuestionado de 1674 hecho por un esclavo fugado no puede ser descartado sin más.[50]
Extinción del solitario de Rodrigues
Aunque la IUCN lista al solitario de Rodrigues extinto para el año 1778,[43] la especie probablemente se extinguió en algún momento entre las décadas de 1730 a 1760; la fecha exacta es desconocida. Su desaparición coincide con la explotación de tortugas entre 1730 a 1750, cuando los comerciantes quemaban la vegetación, cazaban a los solitarios, y liberaron gatos y cerdos que depredaban a sus huevos y polluelos.[51] En 1755, Joseph-François Charpentier de Cossigny trató de conseguir un ejemplar vivo, ya que él estaba seguro de que los solitarios aún sobrevivían en las áreas más remotas de la isla. Tras buscar por 18 meses y ofrecer grandes recompensas, no pudo hallar ninguno. Él señaló que los gatos eran culpados de haber diezmado a la especie, pero sospechaba que en realidad se debía a la caza por los humanos.[51] Cuando Alexandre Guy Pingré visitó Rodrigues para observar el tránsito de Venus de 1761, tampoco encontró a ningún solitario, aunque se le había asegurado que aún existían.[52]
Referencias
Notas
- ↑ Raphidae Poche, 1904 no está disponible porque Poche basó esta familia en el género Raphus Mohring, 1752 el cual no es válido al ser un nombre anterior al sistema de Linneo.
- ↑ La familia Inepti (Illiger, 1811) no está disponible debido a que no fue definida con ningún género.
- ↑ Proviene de un nombre científico hoy obsoleto del dodo, Didus.
- ↑ En 1966, Raphinae fue nombrada para una subfamilia de insectos dentro de Noctuidae. Más tarde se supo que era sinónimo de Dilobinae.
- ↑ El Oxford English Dictionary le da al término original "frontlet" el significado de una banda para la frente o diadema, que es usada en ornitología como la línea de color justo detrás del pico y provee una cita de este uso en 1874.
- ↑ a b Mlíkovsky, 1999.
- ↑ a b c Strickland y Melville, 1848.
- ↑ a b c d e Parish, 2013.
- ↑ Quammen, 1996.
- ↑ a b Sharpe, 1893.
- ↑ Lydekker, 1891.
- ↑ Long, 1843.
- ↑ Reinhardt, 1842–1843.
- ↑ Strickland, 1859.
- ↑ Shapiro et al., 2002 y Janoo, 2005.
- ↑ Hume, Cheke y McOran-Campbell, 2009.
- ↑ a b c Shapiro et al., 2002.
- ↑ BBC, 2002.
- ↑ Owen, 1867.
- ↑ Heupink, Tim H; van Grouw, Hein; Lambert, David M (2014). «The mysterious Spotted Green Pigeon and its relation to the Dodo and its kindred». BMC Evolutionary Biology 14 (1): 136. doi:10.1186/1471-2148-14-136.
- ↑ Pereira et al., 2007.
- ↑ Naish, 2014.
- ↑ Storer, 1970.
- ↑ Janoo, 2005.
- ↑ Mourer-Chauviré et al., 1995.
- ↑ Cheke y Hume, 2008, pp. 70–71.
- ↑ McNab, 1999.
- ↑ Hume, 2012.
- ↑ Fuller, 2001, pp. 37–39.
- ↑ Worthy, 2001.
- ↑ a b c d e Livezey, 1993.
- ↑ Hume, 2006.
- ↑ Fuller, 2003, p. 48.
- ↑ Fuller, 2002, p. 45.
- ↑ a b c Kitchener, August 1993.
- ↑ Angst, Buffetaut y Abourachid, March 2011.
- ↑ Louchart y Mourer-Chauviré, 2011.
- ↑ Angst, Buffetaut y Abourachid, April 2011.
- ↑ Hume y Walters, 2012, pp. 134–136.
- ↑ Brom y Prins, 1989.
- ↑ Rothschild, 1907, pp. 177–179.
- ↑ a b c Fuller, 2001, pp. 203–205.
- ↑ Newton y Newton, 1867.
- ↑ a b c d e Hume y Steel, 2013.
- ↑ Hume y Walters, 2012, pp. 137–138.
- ↑ Rand, 1954.
- ↑ Amadon, 1951.
- ↑ a b c IUCN Red List, 2012b.
- ↑ a b IUCN Red List, 2012a.
- ↑ BBC, 2002-11-20.
- ↑ Hume y Walters, 2012.
- ↑ Fryer, 2002.
- ↑ Cheke y Hume, 2008, p. 79.
- ↑ Roberts, 2013.
- ↑ Roberts y Solow, 2003.
- ↑ a b Cheke y Hume, 2008, pp. 111–114.
- ↑ Fuller, 2002, pp. 156–164.
Bibliografía
- Amadon, D. (1951). «Inbreeding and Disease». Evolution 5 (4): 417. JSTOR 2405692. doi:10.2307/2405692.
- Angst, D.; Buffetaut, E.; Abourachid, A. (2011). «The end of the fat dodo? A new mass estimate for Raphus cucullatus». Naturwissenschaften 98 (3): 233-236. Bibcode:2011NW.....98..233A. PMID 21240603. doi:10.1007/s00114-010-0759-7.
- Angst, D.; Buffetaut, E.; Abourachid, A. (April 2011). «In defence of the slim dodo: A reply to Louchart and Mourer-Chauviré». Naturwissenschaften 98 (4): 359. Bibcode:2011NW.....98..359A. doi:10.1007/s00114-011-0772-5.
- BBC (28 de febrero de 2002). «DNA yields dodo family secrets». BBC News (London). Consultado el 7 de septiembre de 2006.
- BBC (20 de noviembre de 2003). «Scientists pinpoint dodo's demise». BBC News (London). Consultado el 7 de septiembre de 2006.
- BirdLife International (2013). «Pezophaps solitaria». Lista Roja de especies amenazadas de la UICN 2013.2 (en inglés). ISSN 2307-8235. Consultado el 26 de marzo de 2014.
- BirdLife International (2013). «Raphus cucullatus». Lista Roja de especies amenazadas de la UICN 2013.2 (en inglés). ISSN 2307-8235. Consultado el 26 de marzo de 2014.
- Brom, T. G.; Prins, T. G. (June 1989). «Microscopic investigation of feather remains from the head of the Oxford dodo, Raphus cucullatus». Journal of Zoology 218 (2): 233-246. doi:10.1111/j.1469-7998.1989.tb02535.x.
- Cheke, Anthony S. (2004). «The Dodo's last island». Royal Society of Arts and Sciences of Mauritius. Consultado el 12 de mayo de 2012.
- Worthy, T. H. (2001). «A giant flightless pigeon gen. Et sp. Nov. And a new species of Ducula (Aves: Columbidae), from Quaternary deposits in Fiji». Journal of the Royal Society of New Zealand 31 (4): 763-794. doi:10.1080/03014223.2001.9517673.
- Fuller, E.. Extinct Birds. New York: Comstock Pub. pp. 1-398. ISBN 978-0-8014-3954-4. OCLC 46671147.
- Fuller, E. (2002). Dodo: From Extinction to Icon. London: Collins. pp. 1-192. ISBN 978-0-0071-4572-0. OCLC 50877321.
- Fuller, E. (2004). The Dodo: Extinction in Paradise. Lost Worlds. Hawkurst: Bunker Hill Pub. pp. 5-47. ISBN 978-1-5937-3002-4. OCLC 59303394.
- Fryer, J. (2002). «Bringing the dodo back to life». BBC News (London). Consultado el 7 de septiembre de 2006.
- Hume, Julian Pender (2006). «The History of the Dodo Raphus cucullatus and the Penguin of Mauritius». Historical Biology 18 (2): 69-93. ISSN 0891-2963. doi:10.1080/08912960600639400.
- Hume, Julian Pender; Cheke, Anthony S.; McOran-Campbell, A. (2009). «How Owen 'stole' the Dodo: Academic rivalry and disputed rights to a newly-discovered subfossil deposit in nineteenth century Mauritius». Historical Biology 21 (1–2): 33-49. doi:10.1080/08912960903101868.
- Hume, J.P. (2012). Extinct Birds. Poyser Monographs. London: T & AD Poyser. ISBN 978-1-4081-5725-1. OCLC 778339723.
- Hume, J. P. (2012). «The Dodo: From extinction to the fossil record». Geology Today 28 (4): 147-151. doi:10.1111/j.1365-2451.2012.00843.x.
- Hume, J. P.; Steel, L. (2013). «Fight club: A unique weapon in the wing of the solitaire, Pezophaps solitaria (Aves: Columbidae), an extinct flightless bird from Rodrigues, Mascarene Islands». Biological Journal of the Linnean Society: n/a. doi:10.1111/bij.12087.
- Janoo, A. (April–June 2005). «Discovery of Isolated Dodo Bones [Raphus cucullatus (L.), Aves, Columbiformes] from Mauritius Cave Shelters Highlights Human Predation, with a Comment on the Status of the Family Raphidae Wetmore, 1930». Annales de Paléontologie 91 (2): 167-180. doi:10.1016/j.annpal.2004.12.002.
- Kitchener, Andrew C. (August 1993). «Justice at last for the dodo». New Scientist: 24.
- Livezey, B. C. (1993). «An Ecomorphological Review of the Dodo (Raphus cucullatus) and Solitaire (Pezophaps solitaria), Flightless Columbiformes of the Mascarene Islands». Journal of Zoology 230 (2): 247-292. doi:10.1111/j.1469-7998.1993.tb02686.x.
- Long, George, ed. (1833–1843). «The Penny Cyclopaedia of the Society for the Diffusion of Useful Knowledge». Society for the Diffusion of Useful Knowledge (London) 19: 305.
- Louchart, A.; Mourer-Chauviré, C. C. C. (April 2011). «The dodo was not so slim: Leg dimensions and scaling to body mass». Naturwissenschaften 98 (4): 357-358; discussion 358-360. Bibcode:2011NW.....98..357L. PMID 21380621. doi:10.1007/s00114-011-0771-6.
- Lydekker, R. (1891). Catalogue of the Fossil Birds in the British Museum (Natural History). Taylor & Francis. OCLC 4170867. doi:10.5962/bhl.title.8301.
- McNab, B. K. (1999). «On the Comparative Ecological and Evolutionary Significance of Total and Mass-Specific Rates of Metabolism». Physiological and Biochemical Zoology 72 (5): 642-644. JSTOR 10.1086/316701. PMID 10521332. doi:10.1086/316701. }
- Mlíkovsky, J. (1999). «Family-Group Names of Cenozoic Birds: 1811–1998». Paleozoology (Czech Republic: Institute of Geology and Paleontology. Charles University) 169 (1–4): 75-90.
- Mourer-Chauviré, C. C.; Bour, R.; Ribes, S. (1995). «Was the solitaire of Réunion an ibis?». Nature 373 (6515): 568. doi:10.1038/373568a0.
- Naish, D. (2014). «A Review of 'The Dodo and the Solitaire: A Natural History'». Journal of Vertebrate Paleontology 34 (2): 489. doi:10.1080/02724634.2013.803977.
- Newton, Alfred; Newton, Edward (1867). «On the Osteology of the Solitaire or Didine Bird of the Island of Rodriguez, Pezophaps solitaria (Gmel.)». Proceedings of the Royal Society of London 16: 428-433. doi:10.1098/rspl.1867.0091.
- Owen, R. (January 1867). «On the Osteology of the Dodo (Didus ineptus, Linn.)». The Transactions of the Zoological Society of London 6 (2): 49-85. doi:10.1111/j.1096-3642.1867.tb00571.x.
- Parish, J.C. (2013). The Dodo and the Solitaire: A Natural History. Life of the Past. Indiana: Indiana University Press. pp. 1-432. ISBN 978-0-2530-0099-6. OCLC 740630833.
- Pereira, S. L.; Johnson, K. P.; Clayton, D. H.; Baker, A. J. (2007). «Mitochondrial and nuclear DNA sequences support a Cretaceous origin of Columbiformes and a dispersal-driven radiation in the Paleogene». Systematic Biology 56 (4): 656-672. PMID 17661233. doi:10.1080/10635150701549672.
- Quammen, David (1996). The song of the dodo. Island biogeography in an age of extinctions. New York: Scribner. pp. 1-702. ISBN 0684800837. OCLC QH541.5.I8 Q35 1996.
- Rand, A. L. (1954). «On the Spurs on Birds' Wings». The Wilson Bulletin 66 (2): 127-134. JSTOR 4158290. doi:10.2307/4158290.
- Reinhardt, Johannes Theodor (1842–1843). «Nøjere oplysning om det i Kjøbenhavn fundne Drontehoved». Nat. Tidssk. Krøyer. IV: 71-72. 2.
- Roberts, D. L.; Solow, A. R. (November 2003). «Flightless birds: When did the dodo become extinct?». Nature 426 (6964): 245. PMID 14628039. doi:10.1038/426245a.
- Roberts, D. L. (2013). «Refuge-effect hypothesis and the demise of the Dodo». Conservation Biology 27 (6): 1478-1480. PMID 23992554. doi:10.1111/cobi.12134.
- Rothschild, W. (1907). Extinct Birds. London: Hutchinson & Co.
- Shapiro, B.; Sibthorpe, D.; Rambaut, A.; Austin, J.; Wragg, G. M.; Bininda-Emonds, O. R. P.; Lee, P. L. M.; Cooper, A. (2002). «Flight of the Dodo». Science 295 (5560): 1683. PMID 11872833. doi:10.1126/science.295.5560.1683.
- Sharpe, R.B. (1893). Catalogue of the Columbae, or Pigeons, in the British Museum of Natural History. En Sharpe, R.B., ed. «Catalogue of the Birds in the British Museum». Department of Zoology (London: British Museum of Natural History) 21: 628-636. doi:10.5962/bhl.title.8233.
- Storer, R. W. (1970). «Independent Evolution of the Dodo and the Solitaire». The Auk 87 (2): 369-370. JSTOR 4083934. doi:10.2307/4083934.
- Strickland, H.E.; Melville, A. G. (1848). The Dodo and Its Kindred; or the History, Affinities, and Osteology of the Dodo, Solitaire, and Other Extinct Birds of the Islands Mauritius, Rodriguez, and Bourbon. London: Reeve, Benham and Reeve.
- Strickland, H. E. (August 1859). «XVI. On some Bones of Birds allied to the Dodo, in the Collection of the Zoological Society of London». The Transactions of the Zoological Society of London 4 (6): 187-196. doi:10.1111/j.1469-7998.1862.tb08059.x.
- Turvey, S. T.; Cheke, A. S. (2008). «Dead as a dodo: The fortuitous rise to fame of an extinction icon». Historical Biology 20 (2): 149-163. doi:10.1080/08912960802376199.
- Worthy, T. H. (2001). «A giant flightless pigeon gen. Et sp. Nov. And a new species of Ducula (Aves: Columbidae), from Quaternary deposits in Fiji». Journal of the Royal Society of New Zealand 31 (4): 763-794. doi:10.1080/03014223.2001.9517673.
Enlaces externos
 Wikimedia Commons alberga una galería multimedia sobre Raphinae.
Wikimedia Commons alberga una galería multimedia sobre Raphinae.
| Control de autoridades |
|
|---|
 Datos: Q551092
Datos: Q551092- Multimedia: Raphinae / Q551092








