Hemotaksa
Hemotaksa je fenomen u kome telesne ćelije, bakterije, i drugi jednoćelijski ili multićelijski organizmi usmeravaju njihovo kretanje na osnovu određenih hemikalija u njihovoj sredini. Za bakterije je važno da nađu hranu (na primer glukozu) putem plivanja u pravcu najveće koncentracije hranljivog molekula, ili da pobegnu od otrova (na primer fenola).[1] U multicelularnim organizmima, hemotaksa je kritična za ranu (npr. kretanje sperme prema jajetu tokom fertilizacije) i za naknadne faze razvoja (npr. migracija neurona ili limfocita), kao i za normalne funkcije.[2] Osim toga, bilo je uočeno da je mehanizam koji omogućava hemotaksu kod životinja podložan podrivanju u toku metastaze raka.
Hemotaksa se naziva pozitivnom ako se kretanje vrši u pravcu najveće koncentracije hemikalije, i negativna ako je pravac suprotan.
Hemoatraktanti i hemorepelanti
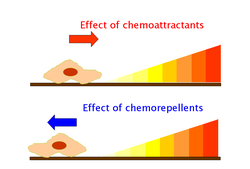
Hemoatraktanti i hemorepelanti su neorganske ili organska supstance koje izazivaju hemotaksu mobilnih ćelija. Hemoatraktanti dejstvuju putem receptora. Oni su specifični ligandi čije efekat je zavisan od koncentracije. Najfrekventnije istraživani hemoatraktanti su formil peptidi i hemokini.[3] Responsi na hemorepelante rezultuju u aksijalnom plivanju i oni se smatraju bazičnim motilnim fenomenom kod bakterija. Najfrekventnije istraživani hemorepelanti su neorganske soli, aminokiseline i neki hemokini.
Bakterijska hemotaksa

Neke bakterije, kao što je E. coli, imaju nekoliko flagela na ćeliji (4–10 tipično). One mogu da rotiraju na dva načina:
- Rotacija nasuprot kazaljki na satu poravnava flagele u jedan rotacioni svežanj, posledica čega je da bakterija pliva u pravoj liniji.
- Rotacija u pravcu kazaljki na satu narušava flagelarni svežanj tako da je svaka flagela usmerena u drugom pravcu, zbog čega bakterija pliva u mestu.
Ponašanje
Sveukupno kretanje bakterija je rezultat naizmeničnih faza kretanja u mestu i plivanja. Ako se posmatra bakterijsko plivanje u uniformnoj sredini, ovo kretanje će izgledati kao randomno kretanje, u kome je relativno pravim plivanjem isprekidano slučajnim prevrtanjem koje reorijentiše bakteriju. Bakterija kao E. coli ne može da izabere direkciju u kojoj će da pliva, i nije u mogućnosti da pliva u pravoj liniji više od nekoliko sekundi usled rotacione difuzije. Drugim rečima, bakterija "zaboravi" pravac u kome se kreće. S obzirom na ta ograničenja, izuzetno je da bakterija može da usmeri svoje kretanje da bi našla povoljnu lokaciju sa visokom koncentracijom atraktanta (obično hrane), i da izbegava repelante (obično otrove).
U prisustvu hemijskog gradijenta bakterija podleže hemotaksi, i usmerava svoje sveukupno kretanje u pravcu gradijenta. Ako bakterija oseti da se kreće u korektnom pravcu (prema atraktantu/nasuprot repelanta), ona će nastaviti da pliva u pravoj liniji duže vreme pre prelaska na kretanje u mestu. Ako se kreće u pogrešnom pravcu, ona će se skorije početi da se kreće u mestu i randomno će probati nov pravac. Drugim rečima, bakterija poput E. coli koristi temporalna očitavanja da odluči da li život postaje bolji ili gori. Na taj način, ona nalazi lokaciju sa najvećom koncentracijom atraktanta (obično izvor) prilično uspešno. Čak i pod veoma visokim koncentracijama, ona još uvek može da pravi razliku između malih koncentracionih razlika. Bežanje od repelanta se odvija sa istom efikasnošću.
Heliksna priroda individualnih flagelarnih filamenata ja kritična za ovo kretanje da se javi. Protein od koga je flagelarni filament formiran, flagelin, je veoma sličan kod svih bakterijskih vrsta koje imaju flagele. Kičmenjaci su iskoristili ovu činjenicu, i razvili imunski receptor (TLR5) koji je dizajniran da prepoznaje ovaj konzervirani protein.
Kao i u mnogim drugim slučajevima u biologiji, postoje bakterije koje ne slede ovo pravilo. Mnoge bakterije, kao što je Vibrio, su monoflagelarne i imaju jedan bič na jednom od polova ćelije. Njihov metod hemotakse je različit. Druge poseduju jedan bič koji se nalazi unutar ćelijskog zida. Te bakterije se kreću vrtenjem u krug cele ćelije, koja ima oblik vadičepa.[4]
Reference
- ↑ George H. Wadhams & Judith P. Armitage (2004). „Making sense of it all: bacterial chemotaxis”. Nature Reviews Molecular Cell Biology 5: 1024–1037. DOI:10.1038/nrm1524.
- ↑ Bruce Alberts, Alexander Johnson, Julian Lewis, Martin Raff, Keith Roberts, Peter Walter (2002). Molecular Biology of the Cell. New York: Garlard Science. ISBN 0-8153-3218-1.
- ↑ Mire-Sluis, Anthony R.; Thorpe, Robin, ur. (1998). Cytokines (Handbook of Immunopharmacology). Boston: Academic Press. ISBN 0-12-498340-5.
- ↑ Berg, Howard C. (2003). „E. coli in motion”. Springer-Verlag, NY. (New York: Springer) ISBN 0-387-00888-8. ISBN 0387008888.
Spoljašnje veze
Hemotaksa na Wikimedijinoj ostavi
- Hemotaksa (knjga)
- Hemotaksa Arhivirano 2014-07-30 na Wayback Machine-u
- Neutrofilna hemotaksa
- Ćelijska migracija Arhivirano 2014-12-11 na Wayback Machine-u
- Globalno postojanje hemotakse sa konačnim radijusom sampliranja Arhivirano 2007-09-27 na Wayback Machine-u
- bakterijska hemotaksa
- Simulacija hemotakse
- Bacterial Interaktivni simulator hemotakse
- p
- r
- u
| |||||||||||||||
| |||||||||||||||
| |||||||||||||||
Monokin • Limfokin (Limfotoksin, Faktor transfera) • Faktor rasta • Hematopoeza (Faktor stem ćelija, Faktor stimulacije kolonije) • Faktor autokrinog motiliteta • Osteopontin
B trdu: peptidi (nrpl/grfl/cytl/horl), receptori (lgic, enzr, gprc, igsr, intg, nrpr/grfr/cytr), itra (adap, gbpr, mapk), calc, lipd, signalni putevi (hedp, wntp, tgfp+mapp, notp, jakp, fsap, hipp, tlrp)






